质体定义2025年03月17日 | 阅读 9 分钟 叶绿体 (plastid) 是一种存在于植物、藻类和某些其他真核生物细胞中的膜结合细胞器(源自希腊语 plast�s,意为“形成、塑造”;复数形式为“plastids”)。它们被认为是细胞内的内共生蓝细菌。例子包括质体(leucoplasts),它们是无色的质体,有时具有分化能力;叶绿体(chloroplasts),用于光合作用;以及有色体(chromoplasts),用于色素的产生和储存。 大约在15亿年前,一个导致陆地植物、红藻和绿藻的古植物类群(Archaeplastida)永久性内共生事件,可能涉及与 Gloeomargarita 属相关的共生蓝细菌(cyanobiont)。之后,在9000万到1.4亿年前,光合保琳氏变形虫(Paulinella amoeboid)发生了一次主要的内共生事件。这种叶绿体属于蓝细菌 Prochlorococcus 和 Synechococcus 的“PS-clade”类群。此外,一些物种通过一种称为“盗食质体”(kleptoplasty)的过程摄取并保留了被吞噬的叶绿体,并且在多种类群中也观察到了次级和三级内共生。 “质体”(plastids)一词及其精确定义最初由 A. F. W. Schimper 提出。叶绿体中含有的色素种类决定了细胞的颜色,并且它们通常包含光合作用所需的色素。它们也是自养真核生物细胞使用的重要化学物质的产生和储存场所。它们拥有一个与原核生物圆形染色体相似的圆形双链 DNA 分子。即使在失去光合能力后,叶绿体仍然被保留,因为它们在异戊二烯类化合物的合成中起着至关重要的作用。 起源质体被认为是内共生蓝细菌。由于大约15亿年前古植物类群的主要内共生事件,真核生物才能够进行产氧光合作用。自那时以来,古植物类群出现了三个进化谱系,每个谱系对叶绿体都有独特的称呼:绿藻或植物中的叶绿体(chloroplasts),红藻中的红质体(rhodoplasts),以及蓝绿藻中的蓝绿藻体(muroplasts)。叶绿体的颜色和超微结构不同。例如,植物和绿藻的叶绿体不再包含蓝细菌、红藻和蓝绿藻中存在的光捕获结构——藻蓝蛋白体(phycobilisomes)。取而代之的是,它们现在只拥有基质和基粒类囊体。与叶绿体和红质体不同,蓝绿藻的叶绿体仍然被蓝细菌细胞壁包裹。这些主要的叶绿体都由两层膜包围。 这是除古植物类群外,唯一已知的蓝细菌主要内共生事件。光合保琳氏(Paulinella)物种的叶绿体,通常被称为“氰体”(cyanelle)或“色素体”(chromatophore),在约9000万至1.4亿年前经历了一次相当晚近的内共生事件。这种叶绿体属于“PS-clade”(属于蓝细菌 Prochlorococcus 和 Synechococcus 属),这与包含古植物类群叶绿体的姊妹谱系不同。 与起源于蓝细菌原初内共生的初生叶绿体不同,复合叶绿体起源于次级内共生,即一个真核生物吞噬了另一个具有初生叶绿体的真核生物。当一个真核生物摄入红藻或绿藻并将其保留时,藻类叶绿体通常被两层以上的膜包围。在各种情况下,这些叶绿体的代谢和光合活性能力可能会减弱。大多数裸藻(红质体)、涡鞭毛藻、隐藻和褐藻等藻类都拥有通过红藻次级内共生产生的复合叶绿体。而眼虫和绿藻虫(叶绿体)等则内共生了绿藻。顶复门(Apicomplexa)是一个专性寄生原生动物类群,其成员包括引起人类或动物疾病的疟原虫(Plasmodium spp.)、弓形虫(Toxoplasma gondii)以及许多其他病原体,它们也都包含复合叶绿体(尽管某些顶复门生物,如引起隐孢子虫病的隐孢子虫 Cryptosporidium parvum,已经丢失了该细胞器)。尽管“顶复质体”(apicoplast)已无法进行光合作用,但它仍然是一个至关重要的细胞器,也是开发抗寄生虫药物的有希望的目标。 一些海蛞蝓和涡鞭毛虫(尤其是 Elysia 属)以藻类为食,并保留被吞噬的藻类叶绿体以从中获得光合作用的好处;最终,叶绿体本身也会被消化。这个过程源自希腊语 kleptes,意为“小偷”,被称为盗食质体(kleptoplasty)。 叶绿体类型根据其颜色、形状和超微结构,叶绿体可分为多种类型。每种叶绿体的性质都与其发挥的功能密切相关。“原叶体”(proplastids)是指未分化的叶绿体,主要存在于分生组织和生殖组织中。它们通常很小,具有独特的超微结构,从而与其他叶绿体类型区分开来。它们可以根据颜色区分,例如“质体”(leucoplasts)为白色,“叶绿体”(chloroplasts)为绿色,而“有色体”(chromoplasts)为黄色、橙色或红色。“绿体”(etioplasts)是指中间形式的叶绿体,而“衰老质体”(gerontoplasts)是指衰老的叶绿体形式。质体(Leucoplasts)因缺乏颜色而区分。然而,它们也可以根据其内容物进一步细分,例如“淀粉质体”(amyloplasts)富含淀粉,“蛋白质体”(proteinoplasts)富含蛋白质,“脂质体”(elaioplasts)富含脂质。 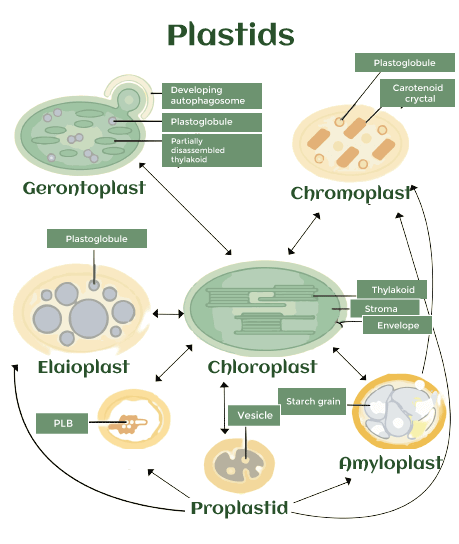 1. 原叶体未分化的叶绿体,称为原叶体,保留了基本的叶绿体结构,用于代际细胞器传递。与其它类型的叶绿体相比,它们是无色的,非常小,并且缺乏任何明显的形态特征。它们主要发现在植物的分生组织和卵细胞中,有时也在某些物种(如 Pelagonium 和大麦)的花粉生成过程中发现。此外,有研究表明,根组织中的根瘤原叶体对于豆科植物的固氮生化过程至关重要。 2. 叶绿体所有进行光合生长的生物都含有叶绿体,这是研究最广泛的叶绿体类型之一。通过使用光合蛋白复合物,它们可以将光能转化为化学能。在叶绿体中,由许多类似圆盘状的类囊体堆叠而成的大型脂质层称为基粒(grana),类囊体用于附着光合蛋白复合物。类囊体边缘形成的称为质体球(plastoglobules)的独特疏水性囊泡结构,有助于增加脂质双层的内表面积。质体球被认为是类胡萝卜素降解的场所,用于产生类胡萝卜素和积累非内源性类胡萝卜素。 叶绿素是光合作用的关键组成部分,赋予叶绿体其特有的绿色。然而,叶绿体还含有大量的其他疏水性萜类化合物,使光合作用得以进行,包括叶黄素、β-胡萝卜素、紫黄质和新黄质。叶绿体中的类胡萝卜素不仅将紫外-蓝光转化为光合作用所需的电能,还通过控制过量激发能的非辐射耗散,显著促进光保护。特别是羟基类胡萝卜素,也称为叶黄素,通过直接淬灭光合作用产生的叶绿素(Chl)的三线态,或清除产生的活性氧(ROS),来帮助光保护。因此,叶绿体的关键作用之一是维持光合作用、光保护和活性氧清除之间的平衡。为了暂时储存并帮助响应发育和环境信号,叶绿体经常生成淀粉颗粒、蛋白质体和脂质体。  3. 绿体绿体是一些中间类型的叶绿体,主要存在于在黑暗中发育的幼苗中。它们很容易在幼苗从土壤中自然萌发时被发现。由于它们会停止合成黑暗中不必要的合成光合作用化学物质和结构,因此它们也被认为是叶绿体的临时发育阶段。在绿体中,通常会形成一个排列良好的、呈多晶体结构的类球状体(prolamellar body)和管状前类囊体(prothylakoids)。这些结构与含有大量类胡萝卜素(特别是叶黄素和紫黄质)的许多小型质体球(plastoglobules)交织在一起,这些类胡萝卜素有助于加速向叶绿体的转变。 质体及其衍生物 构成质体的白色结构非常独特。它们通常存在于与光合作用无关的储存需求组织中。由于显微镜技术的进步和检测方法的增加,质体现在可以进行更深入的分类。三种白色的质体——淀粉质体、蛋白石体和脂质体——除了未分化的原叶体外,被进一步归类为质体的亚型。 淀粉质体 淀粉质体以高密度储存淀粉的淀粉颗粒为特征。在淀粉质体膜形成过程中,淀粉颗粒中存在多种脂质,包括游离脂肪酸、溶血磷脂、溶血磷脂酰胆碱和溶血磷脂酰乙醇胺。淀粉质体通常在各种组织中以低频率存在,包括叶、茎和根,用于暂时储存。它们也经常存在于库组织(sink tissues)中,如种子、果实、块茎和根,用于碳储存。有趣的是,淀粉质体并非总是稳定的;例如,在拟南芥的叶片中,淀粉的积累和丢失非常动态,作为光合作用活动或其缺失的结果,在一个日常周期内发生。淀粉质体通常与其他形式的叶绿体共存于细胞中,这与其他类型的叶绿体不同。在某些植物(如冬南瓜、桃子棕果和红薯块茎)的组织中发现了淀粉质体(amylochromoplasts),这是一种混合型叶绿体,在同一叶绿体中同时储存淀粉颗粒和类胡萝卜素晶体。淀粉颗粒也可能存在于其他类型的叶绿体中,如叶绿体。据报道,拟南芥根部的淀粉质体除了储存功能外,还有助于向地性信号传导。 蛋白石体 在叶绿体结构中——通常在细胞质区域——存在的特定蛋白质体实例被称为蛋白石体。在叶绿体发育的各个阶段,蛋白石体通常可以在多种类型的细胞中发现。由于其位置和组成,人们认为蛋白石体在蛋白质储存中起作用。此外,烟草根部的蛋白石体显示出显著的氧化酶活性,表明它们可能具有特定功能。 有色体 由于储存大量类胡萝卜素,有色体呈现出多种颜色,每种颜色都由不同种类的类胡萝卜素引起。在有色体生长过程中,在疏水性结构质体球(plastoglobules)中产生的和储存的浓缩类胡萝卜素会形成球状、圆形、螺旋状的类胡萝卜素晶体。质体球是与类囊体通过半脂质双层连接的脂蛋白颗粒,它们负责脂质的生产、储存和裂解过程。在生殖组织中,这些具有高度发达质体球的彩色叶绿体被用来吸引传粉者和种子传播者,或用于储存类胡萝卜素和疏水性化合物。 4. 衰老质体衰老质体是为回收而改造的叶绿体,通常在衰老过程或胁迫条件下观察到。叶绿体的分解及其营养物质的回收对植物生命至关重要,因为它们包含植物体内高达80%的氮。此外,据报道,自噬、衰老相关液泡(SAV)和叶绿体囊泡化(CV)都参与了叶绿体蛋白的降解。一旦衰老过程开始,叶绿体会经历一系列超微结构变化。衰老初期衰老质体的性质很难确定。然而,已经注意到了一些独特的特征。首先,衰老质体缺乏淀粉颗粒,可能是因为它们无法维持正常补充淀粉的光合作用。其次是叶绿素和类囊体结构的降解。第三,它们的质体球的大小和数量正在增加,这很可能是由于分解脂质结构和疏水性成分产生的亲脂性物质的积累造成的。 继承在植物中,叶绿体通常只来自一个亲本。大多数裸子植物从雄性花粉获取叶绿体,而被子植物通常从雌配子获取叶绿体。此外,藻类也只从一个亲本接收叶绿体。因此,另一个亲本的叶绿体 DNA 会完全丢失。 在正常的同种杂交(产生正常的同一种属杂种)中,叶绿体 DNA 的遗传呈紧密的一亲遗传。然而,在种间杂交中,叶绿体的遗传更加不规律。尽管种间杂交中的叶绿体通常是母系遗传的,但在一些开花植物杂种中,也存在父系特异性叶绿体的情况。苜蓿(Medicago sativa)在20%的被子植物中,通常表现出双亲叶绿体遗传。 DNA 的修复与损伤正在生长的玉米幼苗的叶绿体 DNA 更容易受到损伤。DNA 在光氧化反应和光合/呼吸电子传递产生的氧化条件下会受到损伤。虽然一些 DNA 分子会得到修复,但未修复的 DNA 损伤会导致其降解成无功能片段。 DNA 修复蛋白由细胞的核基因组编码。但它们可以被转运到叶绿体,在那里修复叶绿体 DNA 以确保基因组的稳定/完整性。例如,为了保持叶绿体基因组的完整性,DNA 错配修复蛋白 Msh1 与重组修复蛋白 RecA 和 RecG 在苔藓 Physcomitrella patens 的叶绿体中相互作用。 总结与展望对多个代表性样本的分析表明,叶绿体相互转化的复杂调控是许多农艺性状的关键组成部分。叶绿体的发育和相互转化不仅影响了淀粉、脂肪、蛋白质和次级代谢产物等多种植物源产品的生产潜力。尽管面临环境条件的限制和不断增长的植物需求,研究叶绿体可能是在寻找突破性解决方案的关键。它还为叶绿体研究提供了科学证据,包括作为分子修饰候选对象的核心调控因子。最后,通过简要描述的叶绿体转化分子通路获得的候选基因,可以作为目标基因,以增强与叶绿体转化相关的各种农艺性状。 下一主题聚合定义 |
我们请求您订阅我们的新闻通讯以获取最新更新。
我们提供所有技术(如 Java 教程、Android、Java 框架)的教程和面试问题
G-13, 2nd Floor, Sec-3, Noida, UP, 201301, India
